Le Contexte historique et démographique de la population de Honaïne
La région de Honaïne, grâce à son passé historique et culturel, s’inscrit bien dans l’héritage du patrimoine architectural riche et diversifié du chef lieu de la wilaya de Tlemcen. Au niveau ethnique, cette population appartient à la tribu des Koumia établi dans les monts de Taras avant le XVIème siècle. La population berbère de Honaïne est une population endogame à faible effectif, sa structure et son relief géographique feraient d’elle une population de référence (Aouar et al., 2005 ; Otmani et al., 2017), qui nous incitera à évaluer sa diversité génétique et la comparer d’une part aux populations inter-régionales et d’autre part aux populations de l’espace Méditerranéen. La figure 01 présente l’arbre d’organisation tribale de la population de Honaïne.
Honaïne dans l’antiquité
Au début, la ville a été fondée en tant que comptoir commercial dont les jalons ont été posés par les commerçants phéniciens aux environs du Xllème siècle avant Jésus Christ (J.C.) (Lethielleux, 1974) à l’instar des comptoirs du rivage méridional méditerranéen. Elle est connue sous le nom GYPSARIA PORTUS (Mc Carthy, 1956), le mot désignant le lieu du gypse dont regorgent les cimes élevées lequel varie du calcaire transformé en marbre sous diverses formes naturelles. Sous l’ère de l’occupation romaine, la ville paraît plus moderne afin d’aller de pair avec les exigences des fonctions qui lui ont été dévolues, étant donné qu’il a été procédé à la réparation du port et à son extension à la terre ou un bassin de dimension (85m/50m) a été creusé, en étant relié par un canal au port (Khalifa, 1988).
Aspect démographique
Honaïne est une daïra composéede deux communes qui comptent 12495 habitantsau 31 décembre 2011 (commune de Honaïne, et la commune de Beni Khellad) (Monographie de Tlemcen, 2011). Dans la commune de Honaïne, les urbains représentent environ 82.5% de la population totale. D’où le taux de la population rurale n’est quede 27.5% (PDAU, 2005). Le nombre d’habitants de la commune de Honaïne est de 5402 soit une densitéde 95 Habitants/Km2 . Lenombre d’habitants de la commune de Beni Khellad est 7093 habitants soit une densité de 83 Habitants/Km2 (Tableau 01).
Paramètres d’étude de la diversité génétique
La génétique des populations humaines s’intéresse aux mécanismes et aux causes de l’évolution des fréquences alléliques au cours du temps au sein des populations et à la diversité génétique de ces dernières (Cavalli-Sforza, 1998 ; Harry, 2008). Pendant des décennies, les os fossiles et les témoignages archéologiques ont été les seuls outils pour reconstituer l’histoire de la lignée humaine. Depuis une vingtaine d’année, l’étude de l’ADN des populations contemporaines constitue une source supplémentaire de connaissances (Radin, 2015). Les résultats génétiques obtenus doivent, être confrontés aux approches historiques, paléontologiques, archéologiques, paléo-climatologiques et linguistiques (Bauduer, 2013). La génétique des populations a des applications en épidémiologie où elle permet de comprendre la transmission des maladies génétiques (Bonello et al., 2009).
Les marqueurs érythrocytaires et la notion de groupes sanguins
Le sang est considéré comme l’un des outils essentiels pour comprendre l’origine ethnique des personnes et pour la transfusion sanguine. Les groupes sanguins ont un mécanisme héréditaire distinct qui permet de mesurer la variabilité biologique en termes de caractères car ils ne sont pas soumis à l’influence de l’environnement (Ara et al., 2011). Les groupes sanguins sont des marqueurs génétiques présentant un grand degré de polymorphisme, ce qui leur donne un intérêt particulier dans les études de micro différentiation et de l’histoire migratoire des populations (Lefevre et Witier 2006 ; El Ossmani, 2008 ; Clavier, 2012).
Le polymorphisme immunogène regroupe des antigènes portés par des cellules [érythrocytaires : groupes sanguins érythrocytaires, leucocytaires : groupes sanguins leucocytaires, plaquettaires : groupes sanguins plaquettaires, toutes cellules nucléées : groupe HLA] ou par des molécules libres dans le torrent circulatoire [immunoglobulines : marqueurs Gm, Am, Km, protéines du complément : groupes Chido Rodgers] (Chiaroni, 2003 ; Neffati et al., 2019) (Figure 02). Son exploration est basée sur la mise en œuvre de techniques immunologiques : utilisation d’anticorps ; si l’anticorps se fixe, c’est que l’antigène est là et donc que l’allèle correspondant est présent. Son champ d’application intéresse surtout le médecin puisque le polymorphisme immunogène s’oppose à la transfusion, la greffe, la transplantation et même la grossesse incompatible. De même, ces marqueurs peuvent être la cible du propre système immunitaire de l’individu, base des pathologies auto-immunes (Christian et Mannessier, 2002).
Quatre systèmes distincts (ABO, Rhésus, MNSs et Duffy) ont été étudiés par des méthodes immunologiques. Ces systèmes ont été sélectionnés pour trois raisons ; (i) leur capacité, surtout lorsqu’ils sont cumulés, à définir différentes populations humaines, (ii) pour les nombreuses données bibliographiques existant dans de multiples populations et enfin (iii) pour l’existence possible d’une pression sélective sur certains d’entre eux, notamment pour le système Duffy (Hamblin et al., 2002 ; Hamblin et Di Rienzo, 2000). En effet, ce mécanisme semble impliqué dans l’apparition de variants caractéristiques de régions particulières comme les zones impaludées à Plasmodium vivax d’Afrique subsaharienne, ce qui les fait considérer comme des marqueurs pertinents pour l’étude des migrations des populations humaines.
Le système ABO
Le système de groupe sanguin ABO est sans aucun doute le plus important du fait de son implication en transfusion sanguine et pour la transplantation d’organe. Il présente deux caractéristiques qui sont à l’origine des méthodes de groupage sanguin et expliquent son rôle important en transfusion et transplantation :
• la présence constante d’anticorps « naturels » anti-A et anti-B correspondants aux antigènes absents des globules rouges.
• les antigènes ABO ne sont pas de simples antigènes de groupe sanguin érythrocytaire mais leur présence dans la plupart des tissus et liquides biologiques en fait de véritables antigènes tissulaires (Janot et Mannessier, 2002 ; Lefrère et Rouger, 2009) .
La présence ou l’absence des antigènes A ou B à la surface des hématies est sous la dépendance de trois gènes:
– le gène A qui induit la synthèse de l’antigène A,
– le gène B qui induit celle de l’antigène B,
– le gène O qui ne modifie pas la substance de base H.
La transmission héréditaire de ces gènes obéit aux lois de Mendel. Ces gènes A, B et O sont des allèles, ils sont situés sur un même locus chromosomique et ils s’excluent toujours lors de la méiose. Le groupe sanguin de tout individu dépend donc de la présence de deux de ces trois allèles A, B et O. Les gènes A et B sont codominants, ils s’expriment au niveau du phénotype. Le gène O est quant à lui « récessif » par rapport aux gènes A et B. Ainsi aux phénotypes O et AB, correspond un seul génotype. Pour les groupes A et B, le phénotype peut être différent du génotype. En effet, si les hématies d’un individu sont agglutinées par l’anticorps anti-A et lui seul, elles sont dites de groupe A mais cela n’indique pas si au niveau du génome, il y a deux gènes A ou simplement un gène A associé à un gène O.
✔ Distribution populationnelle
L’étude des groupes ABO, pour des besoins transfusionnels, démontra très tôt que leur répartition variait en fonction des peuples. Certains auteurs ont liés la distribution mondiale du polymorphisme du système ABO à des grandes épidémies et à certaines maladies infectieuses. Ainsi, la fréquence élevée de l’allèle ABO*O chez les Amérindiens peut être attribuée à un avantage sélectif de cet allèle pour la réponse immunitaire à la Syphilis. La fréquence relativement élevée de l’allèle ABO*B chez les populations Asiatiques peuvent être le résultat d’une double action sélective de la peste contre l’allèle ABO*O et de la variole contre l’allèle ABO*A (Vogel et Motulski, 1982). La fréquence de l’allèle A est élevée en Europe surtout dans la région Scandinave et dans certains régions de l’Europe centrale ; des fréquences élevées sont aussi élevés chez les Aborigènes de l’Australie du sud ouest, mais c’est chez certains tribus indiennes de l’ouest d’Amérique du nord que l’on rencontre la fréquence la plus élevée (Goudemand et Salmon, 1980). Malgré tout, à l’exception des Amérindiens, les fréquences des groupes ABO sont assez constantes dans le monde et varient moins d’un endroit à l’autre que d’autres gènes (CavalliSforza et al ., 1994).
Le Système Rhésus
Le système RH représente une belle illustration du polymorphisme. C’est le système de groupe sanguin le plus complexe. Les antigènes Rh sont codés par deux gènes homologues étroitement liés sur le bras court du chromosome 1. Le gène RHD est présent chez 85 % des sujets d’origine européenne qui sont dits RH1. Il est absent ou non fonctionnel chez 15 % des sujets dits RH: -1. Le gène RHD produit la protéine RhD porteuse de l’antigène D. Le gène RHCE produit la protéine RhCE porteuse, en fonction des formes alléliques, des antigènes C ou c et E ou e (RH2, RH4, RH3, RH5) (Wagner et al., 2005 ). D’un point de vue fonctionnel, cette molécule paraît plus particulièrement impliquée dans des fonctions de transport transmembranaire. Compte tenu de l’absence de recombinaison, les deux gènes, donc les trois antigènes, sont transmis en bloc sous forme d’haplotypes qui sont notés DCe, DcE, dce, Dce, dCe, dcE, DCE et dCE où d représente l’allèle RHD en délétion ou inactif.
|
Table des matières
Introduction générale
1. Synthèse Bibliographique
1.1 Le Contexte historique et démographique de la population de Honaïne
1.1.1 Honaïne dans l’antiquité
1.1.2 Honaïne dans le médiéval
1.1.3 Honaïne dans le contemporain
1.1.4 Aspect démographique
1.2. Paramètres d’étude de la diversité génétique
1.2.1 Les marqueurs érythrocytaires et la notion de groupes sanguins
1.2.1.1 Le système ABO
1.2.1.2 Le système Rhésus
1.2.1.3 Le système Duffy
1.2.1.4 Le système MNSs
1.2.2 Croisements non aléatoires (Endogamie et Consanguinité)
1.2.2.1 Endogamie
1.2.2.2 Consanguinité
1.2.2.3 L’impact de la consanguinité sur la morbidité et la mortalité
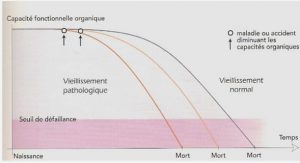
1.2.3 Marqueurs de Morbidité
1.2.3.1 Généralités
1.2.3.2 La morbidité cancéreuse
1.2.3.3 Le profil épidémiologique des cancers dans le contexte Algérien
2. Matériels et Méthodes
2.1 Présentation générale de la population endogame de Honaïne
2.2 Analyses des d’échantillons par type du marqueur
2.2.1 Les marqueurs érythrocytaires : les groupes sanguins
2.2.1.1 Echantillonnage
2.2.1.2 Prélèvement du sang
2.2.1.3 Groupage sanguin
2.2.1.4 Extraction de l’ADN
2.2.1.5 Analyses statistiques
2.2.2 Consanguinité
2.2.2.1 Questionnaire
2.2.2.2 Recrutement des individus
2.2.3 Les marqueurs de morbidité : Le Cancer
2.2.3.1 Structures sanitaires
2.2.3.2 Type d’étude
2.2.3.3 Recueil des données
2.2.3.4 Le traitement statistique des données
3. Résultats et Discussion
3.1 Le polymorphisme érythrocytaire
3.1.1 Fréquences phénotypiques et alléliques des systèmes sanguins
3.1.1.1 Système ABO
3.1.1.2 Système Rhésus
3.1.1.3 Système MNSs
3.1.1.4 Système Duffy
3.1.1.5 Hétérozygotie
3.1.2 Comparaisons inter-populationnelles
3.1.3 La diversité génétique
3.1.4 Affinités inter-populationnelles
3.1.4.1 Analyse en Composante Principales (ACP)
3.1.4.2 Distances et arbres phylogénétiques
3.2 Consanguinité
3.2.1 Fréquence et nature de la consanguinité
3.2.2 Comparaison inter-populationnelle du taux de consanguinité à Honaïne
3.2.2.1 Variabilité inter-circonscriptions
3.2.2.2 Dans le monde Arabo-musulmans
3.3 Marqueurs de morbidité (La morbidité cancéreuse)
3.3.1 Répartition globale du cancer tous types confondus dans la population endogame de Honaïne
3.3.1.1 Prévalence et incidence du cancer
3.3.1.2 Répartition de taux d’incidence du cancer par sexe
3.3.1.3 Répartition de taux d’incidence du cancer par tranche d’âge
3.3.1.4 Répartition de taux d’incidence du cancer par tranche d’âge et par sexe
3.3.2 Variabilité inter-circonscriptions du taux d’incidence du cancer tous types confondus
3.3.2.1 Répartition de taux d’incidence du cancer par circonscriptions
3.3.2.2 Répartition de taux d’incidence du cancer par circonscriptions et par sexe
3.3.2.3 Répartition de l’incidence du cancer par circonscriptions et par tranche d’âge
3.3.3 Evolution chronologique du cancer
3.3.4 Variabilité inter-circonscriptions par type du cancer
3.3.4.1 Variabilité inter-circonscriptions de taux d’incidence du cancer
3.3.4.2 Variabilité inter-circonscriptions de la fréquence du cancer
3.3.5 Variabilités inter-populationnelles de taux d’incidence du cancer en Algérie
3.3.6 Variabilités inter-populationnelles du taux d’incidence du cancer dans le bassin méditerranéen
3.3.6.1 Le cancer du sein
3.3.6.2 Le cancer du poumon
3.3.6.3 Le cancer du cerveau
3.3.6.4 Le cancer du lymphome
3.3.6.5 Le cancer de bouche- pharynx
3.3.7 Les affinités inter-populationnelles
3.3.7.1 Affinité inter-circonscription
3.3.7.2 Affinité inter-populationnelles au niveau national
3.3.7.3 Affinité inter-populationnelles au niveau du bassin méditerranéen
Conclusion
![]() Télécharger le rapport complet
Télécharger le rapport complet