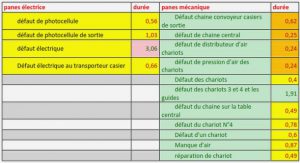
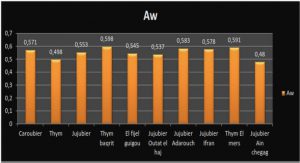
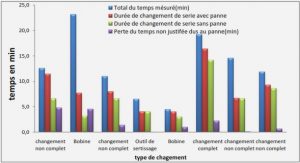
Applications of CBMs
In recent years, CBMs have been used for several applications related to biomedicine, environment, molecular biology, microarrays, paper, textile, food and biofuel industries [142] (Figure 1.13). There are three basic properties of CBMs which contributes to their compatibility for the applications mentioned above: (1) CBMs are independent folding modules and can function autonomously in a chimeric protein; (2) CBMs are diverse, abundant, highly substrate specific and inexpensive (in terms of their production ةand purification as both wild type and recombinant proteins) with excellent chernical and physical properties; and (3) the binding specificities and affinities of CBMs can be, in principle, controlled according to an existing problem (using directed evolution for instance).
Bloprocesslng
One of the major applications ofCBMs is in bioprocessing ofbiologically activemolecules for various biotechnological applications. CBMs have been studied and used as high-capacity purification tags for the isolation, production and purification of biologically active target peptides and proteins at relatively low cost. Numerous other reports have also shown CBM as an affinity tag for enzyme immobilization and processing. Another area of CBMs bioprocessing application includes enhanced bioethanol production from cellulosic materials. A study by Berdichevsky et al. showed that a CBM could be used for matrix-assisted refolding of a single-chain antibody in order to prevent the aggregation of protein during the course of renaturation [143]. In another study, a CBM fused to single chain antibody increased the efficiency of the phage display screening process for recombinant antibodies [144].
Targeting
Cellulose is the most abundant and renewable substrate with various commercial applications. Therefore, CBMs can be used for targeting desired functional molecules to the substrate containing cellulosic fibers, such as cotton fibers in the textile industries. CBMs have been used extensively for the denim washing, an alternative to the original abrasive stonewashing and laundry powders for targeting recombinant enzymes (amylases, proteases and lipases) and fragrance particles to the cellulosic fibers [145,146].
Cell immobilization
CBMs have been used efficiently for the cell immobilization technology for various applications including ethanol production and phenol degradation. CBMs were also used for immobilization of mammalian cells to a cellulosic surface for improving the performance ofvascular graft [147] and cartilage regeneration [148].
Bioremediatlon
CBMs have been studied and shown to be an excellent mediator for the bioremediation of the pollutants. They have been used as recombinant proteins for the removal of toxic heavy metals such as cadmium [149] and a commonly used pesticide atrazine [150] from water. These findings have shown strong potential for the improvement of sustainable wastewater treatment technologies [151].
Analytlcal tool
Many studies have been using CBMs as analytical tools for research and diagnostics of cellulosic fiber surface. McCartney et al. [152] and Jamal-Talabani et al. [153] developed novel molecular CBM probes for the detection, characterization and mapping of plant cell wall polysaccharides. The CBMs were also used in the optimization of the bioprocesses such as fermentation [154], rapid detection of the pathogenic microbes in food samples [155] and production ofCBM based non-DNA microarrays for various medical applications [156].
Modification of fibers
The CBMs have also been used for the modification of cellulosic fibers in the paper and textile industries [157-159]. A recombinant CBM3 conjugated with polyethylene glycol (PEG) has shown to improve the drainability of the wood pulps without affecting the physical properties of the paper sheets [160]. The recombinant CBMs were also applied to Whatman filter paper and it was found to enhance its me’Chanical properties, such as tensile strength, brittleness, Young’s modules and paper dry strength. Moreover, a recent study by Shi et al. (2014) demonstrated that the use of recombinant double CBMs, such as CBMI-NL-CBM1, significantly enhanced the mechanical properties ofpulp and papers [161]. Additionally, CBMs were also successfully used for the scouring process, one of the most important process determining the fabric quality, to remove the cutical layer of cotton fibers and increasing the efficiency of the process [162].
Fluorescent proteln-tagged CBMs
Fluorescence is a very sensitive and specific spectroscopy where absorption and emission wavelengths determine what molecules contribute to the detected signaIs [125,163]. Further, plate readers allow increasing measurement throughput, a valuable criterion in the development of any novel assay. Rence, detection of CBM probes that wou Id emit fluorescence would be advantageous. Fluorescence detection can be achieved directly or indirectly depending on the methods used [118]. The indirect methods involve the use of a secondary or tertiary reagent such as anti-Ris-IgG coupled to a fluorophore to detect the Ris-tag of a CBM, which may also allow amplification of signal intensities.
This method provides great flexibility in CBM use but has a potential disadvantage related to multi-step incubations, which decrease analysis speed and are less compatible with a high throughput strategy [118]. On the other hand, in direct methods, coupled CBMs would require a straightforward, single-step incubation, affording the possibility of rapid, high throughput protocols. In the first direct method reported, a CBM was chemically coupled with a fluorophore (such as FITC/Alexa Fluor) [118]. Unfortunately, these molecules react non-specifically with various moieties at the surface of CBMs, deleteriously affecting specificity, affmity and detection reproducibility. Another direct detection method uses CBMs genetically fused to a fluorescence protein such as the green fluorescent protein (or any of its variants) [118]. This method allows maintenance of the original CBM behavior, avoiding the limitations described for the fIfst direct method discussed. Rence, CBMs coupled with fluorescence protein have been used for mapping the chemistry and structure of various carbohydrate-containing substrates (i.e. LCB) [164,165].
The production and purification ofthese fluorescent protein-tagged carbohydrate-binding modules is part of a broader ‘recombinant CBM-fusion technology’ , which has been used to obtain CBMs fused with other proteins for specifie applications. Recombinant CBMs can either be produced in heterologous organisms (Escherichia coli and Pichia pastoris) or in the CBM’s original host. Figure 1.14 illustrate the recombinant CBM-fusion technology from cloning to applications [121].
Fluorescent protein
Green fluorescent proteln (GFP)
GFP, the famous chemiluminescent protein, was discovered by Shimomura et al. [166] from Aequorea jellyfish. GFP is an 11-stranded ~-barrel structure threaded by an (lhelix running up the axis of the cylinder, which is also known as ~-can (Figure 1.15). The ability of fluorescent proteins to emit visible light derives from the post-translational modification ofthree amino acids at positions 65-67 (Ser-Tyr-Gly in the native protein), which result in chromophore formation. The chromophore, a p-hydroxybenzylideneimidazolinone, is attached to the a-helix and is buried in the center of the cylinder. Figure 1.16 shows the mechanism for chromophore formation. First, GFP folds into a nearly native conformation, then the imidazolinone is formed by nucleophilic attack of the amide of Gly67 on the carbonyl ofresidue 65, followed by dehydration. Finally, molecular oxygen dehydrogenates the (l-~ bond ofresidue 66 to put its aromatic group into conjugation with the imidazolinone. Only at this stage the chromophore acquires visible absorbance and fluorescence. The ~-can structure protects the chromophore and is presumably responsible for GFP’ s stability [167]. Figure 1.17 and Table 1.4 describes the different classes of green fluorescent proteins and their properties [168].
|
Table des matières
Acknowledgements
Abstract
Résumé
Table of Content
List of Figures
List of Tables
List of Additional Files
List of Abbreviations
Chapter 1 – Introduction
1.1 Lignocellulosic sources
1.1.1 Forest woody feedstocks
1.1.2 Agricultural residues, herbaceous and municipal solid wastes
1.1.3 Algae biomass
1.2 Lignocellulosic biomass composition
1.2.1 Cellulose
1.2.2 Hemicellulose
1.2.3 Lignin
1.3 Lignocellulosic biomass recalcitrance
1.3.1 Pretreatments
1.3.2 Enzymatic hydrolysis
1.4 Analytical methods for investigating biomass recalcitrance
1.5 Carbohydrate-binding modules
1.5.1 CBMs classification
1.5.1 .1 Protein fold classification
1.5.1.2 Structural and functional classification
1.5.2 Structural determinants ofCBMs binding specificity
1.5.3 CBMs functions
1.5.4 Applications of CBMs
1.5.4.1 Bioprocessing
1.5.4.2 Targeting
1.5.4.3 Cell immobilization
1.5.4.4 Bioremediation
1.5.4.5 Analytical tool
1.5.4.6 Modification of fibers
1.6 Fluorescent protein-tagged CBMs
1.6.1 Fluorescent prote in
1.6.1.1 Green fluorescent protein (GFP) 1.7 Fluorescent protein-Tagged Carbohydrate-binding module Method (FTCM)
1.7.1 Farnily3 CBM
1. 7.2 Farnily 17 CBM
1.7.3 Farnily 15 CBM
1.7.4 Farnily 27 CBM
1.8 Objectives
1.8.l Objective 1 (scientific article 1)
1.8.2 Objective II (scientific article II)
1.8.3 Objective III (scientific article 111)
1.8.4 Objective IV (scientific article IV)
1.8.5 Objective V (scientific article V)
Chapter 2 – Scientific Article 1
2.1 Abstract
2.2 Introduction
2.3 Materials and methods
2.3.1 Chernicals and strains
2.3.2 Construction ofpET11a-mOrange2-CBM15 expression vector
2.3.3 Expression and purification ofOC15 probe
2.3.4 Affinity gel electrophoresis (AGE)
2.3.5 Isothermal titration calorimetry (ITC)
2.3 .6 Pulp characterization
2.3 .7 Handsheets preparation
2.3.8 Xylanase digestion ofunbleached kraft pulp
2.3.9 X-ray photoelectron spectroscopy (XPS)
2.3.10 X ylan tracking on the surface of papers using the OC 15 probe
2.4 Results and Discussion
2.4.1 OC15 expression and purification
2.4.2 Determination of OC 15 ligand specificity using affinity gel electrophoresis (AGE)
2.4.3 Determination of OC 15 ligand affinity using isotherrnal titration calorimetry (ITC)
2.4.4 Comparing XPS, NREUfP-510-42618 and OC15 methodologies for the detection ofxylan
2.4.5 Monitoring xylan hydrolysis using NRELffP-510-42618, XPS and the OC 15 probe
2.5 Conclusion
2.6 References
2.7 Additional Files
Chapter 3 – Scientific Article II
3.1 Abstract
3.2 Introduction
3.3 Materials and methods
3.3.1 Chemicals and microbial strains
3.3.2 Construction, production and purification of CBM recombinant probes
3.3.3 Affinity gel electrophoresis (AGE)
3.3.4 Isothermal titration calorimetry (ITC)
3.3.5 Pulp characterization
3.3.6 Handsheets preparation
3.3.7 Enzymatic digestion ofpaper dises
3.3.8 Lignocellulosic polymers tracking on the surface ofpaper dises using the OC15, CC27 and GC3a probes
3.4 Results and Discussion
3.4.1 Determination of the CC27 probe specificity using affinity gel electrophoresis (AGE)
3.4.2 Detennination ofCC27 probe affinity using isotherrnal titration calorimetry (ITC)
3.4.3 Tracking hernicelluloses at the surface ofwood biomass
3.4.4 Investigation of reaction parameters by FTCM
3.4.5 Addressing possible impact of enzyme inactivation
3.4.6 Investigating the impact of cellulose on the hydrolysis of hemicelluloses
3.4.7 Exploring the impact ofxylan polymers on the hydrolysis of mannan
3.5 Conclusion
3.6 References
3.7 Additional Files
Chapter 4 – Scientific Article III
4.1 Abstract
4.2 Introduction
4.3 Materials and methods
4.3.1 Reagents and pulps
4.3.2 Pulps Characterization
4.3.3 Pulps refming and paper sheets formation
4.3.4 Enzymatic digestions ofpulps and paper dises
4.3.5 Fiber quality analysis and paper physical properties determination
4.3.6 Construction of the recombinant probe expression systems
4.3 .7 Expression and purification of recombinant probes
4.3.8 Quantification of the variations of the carbohydrates signatures on the surface offiber dises
4.3.9 Detennination of the probes affinity for Avicel and fiber dises
4.4 Results and Discussion
4.5 Conclusions
4.6 References
4.7 Additional Files
Chapter 5 – Scientific Article IV
5.1 Abstract
5.2 Introduction
5.3 Materials and methods
5.3.1 Lignocellulosic biomass
5.3.2 Enzyme solutions
5.3.3 Enzymatic treatments ofpulp
5.3.4 Handsheet and paper disc preparation
5.3.5 Construction of recombinant probe expression systems
5.3.6 Expression and purification of probes
5.3.7 Quantification of the carbohydrates on the surface offiber paper discs using FTCM
5.3.8 Sugar analysis
5.3.9 Scanning electron microscope (SEM) images
5.3 .10 Statistical analysis
5.4 Results and discussion
5.4.1 Enzyme characterization
5.4.2 Pulp fiber characterization
5.4.3 Hydrolysate analysis
5.4.4 Effect of enzymatic treatment on pulp fibers
5.4.5 Detection ofpulp fiber polymers using FTCM analysis before and after enzymatic treatments
5.4.6 Surface polymer distribution after enzymatic treatments
5.5 Conclusion
5.6 References
5.7 Additional Files
Chapter 6 – Scientific Article V
6.1 Abstract
6.2 Introduction
6.3 Materials and methods
6.3.1 Chemicals, microbial strains and LCB
6.3.2 Construction, expression and purification of tluorescent-tagged
carbohydrate-binding module probes
6.3.3 Determination of probes affmities and specificities
6.3.4 LCB preparation and pretreatments
6.3.5 Determination of cellulose, hemicellulose and lignin content
6.3.6 Enzymatic treatment ofLCB
6.3.7 Quantification of the variations of the carbohydrates on the
surface of LCB using FTCM -depletion assay
6.3.8 X-ray diffraction (XRD)
6.4 Results and Discussion
6.4.1 Adaptation ofFTCM to a depletion assay for investigation of biomass suspensions
6.4.2 Tracking surface accessibility of lignocellulosic components in LCB
6.4.3 Tracking surface accessibility of lignocellulosic components in pretreated LCB
6.5 Conclusion
6.6 References
6.7 Additional Files
Chapter 7 – Conclusions
Bibliography
Appendix A
Appendix B
Appendix C
Appendix D
![]() Télécharger le rapport complet
Télécharger le rapport complet